Tutorial¶
A recurrent area cell¶
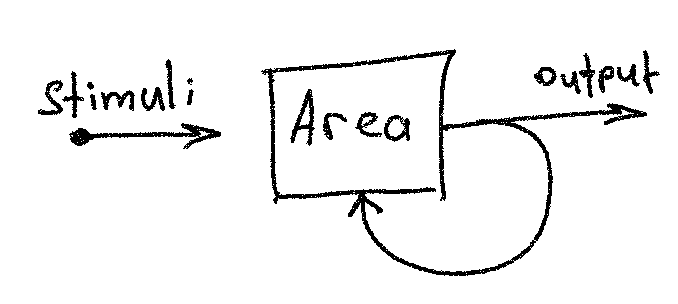
The building block of computation with assemblies 1 is a recurrent neural
network cell, called area, the forward pass of which is described in the
Usage. The output of such an area is a binary sparse vector, formed by
winner-take-all competition. For example, if an input vector multiplied by a
weight yields vector z = [-3.2, 4.6, 0, 0.7, 1.9], then
kwta(z, k=2) = [0, 1, 0, 0, 1].
K-winners-take-all¶
One of the properties of kWTA is that the inverse of kWTA is also kWTA, even
in case of a random projection (the multiplication matrix is random, not
learned). On the plot below, several images from MNIST dataset are shown
on the left, their random projection & cut binary vector
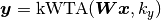 , reshaped as a matrix, in the
middle, and the restored
, reshaped as a matrix, in the
middle, and the restored 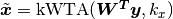 is shown on the right.
is shown on the right.  condition must hold in order to restore the input signal.
condition must hold in order to restore the input signal.

This example shows that a random projection & cut operation (kWTA followed by multiplication by a random matrix) preserves enough information to reconstruct the input signal.
How does the association work?¶
How to associate information from two and more different modalities? For example, how to associate a picture of an elephant with the sound an elephant makes?
Willshaw’s model¶
Let’s define the task in mathematical terms: let x and y denote the image
and the sound representation vectors of a signal respectively. Then the
simplest way to associate x and y is to resort to the Hebbian-like learning
rule. Assuming both x and y are binary sparse vectors, we can construct
the weight matrix as an outer product of x and y. This technique is
described in 2 and implemented in AreaRNNWillshaw.
The idea behind Willshaw’s paper is based on the outer product property:
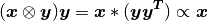
which naturally suggests the following update rule:
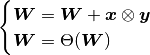
where 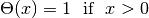 ; otherwise, it’s zero.
; otherwise, it’s zero.
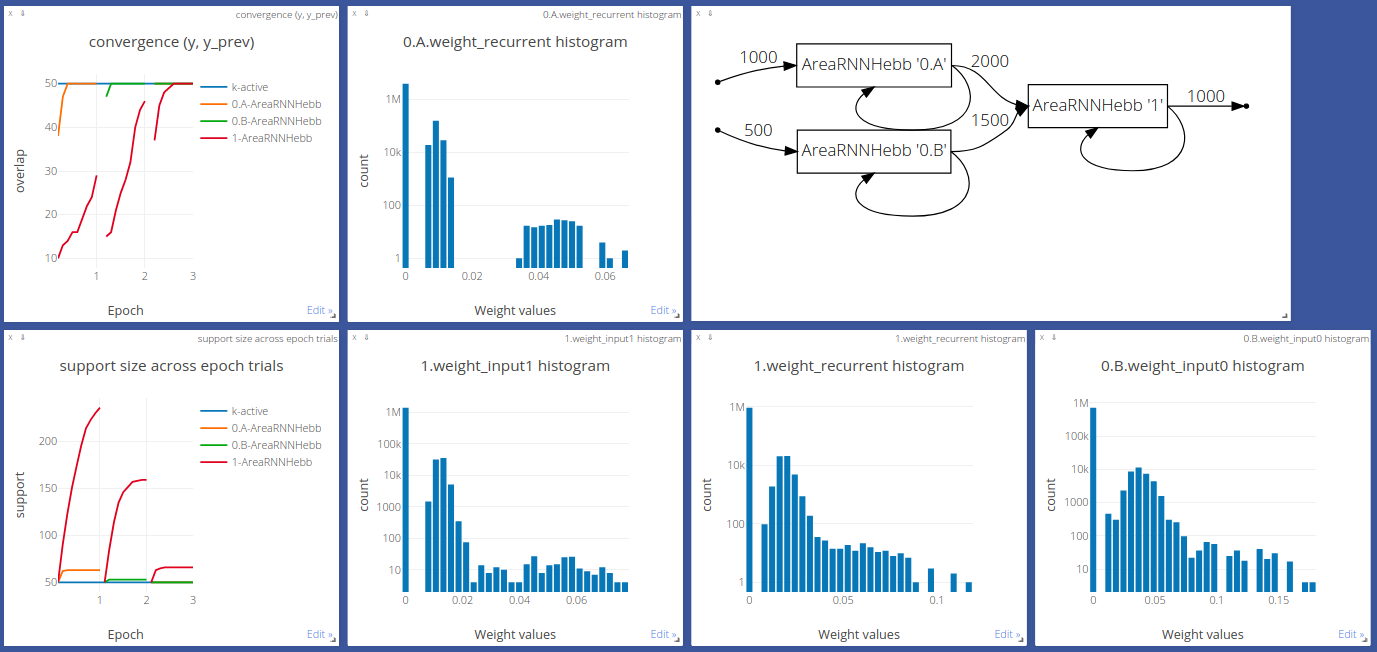
Papadimitriou’s model¶
Willshaw’s update mechanism has a limitation: the initial matrix  must be initialized with zeros, which poses biological plausibility problems.
To alleviate this, we can use a third layer C to indirectly associate the
parental layers A and B, as shown below.
must be initialized with zeros, which poses biological plausibility problems.
To alleviate this, we can use a third layer C to indirectly associate the
parental layers A and B, as shown below.
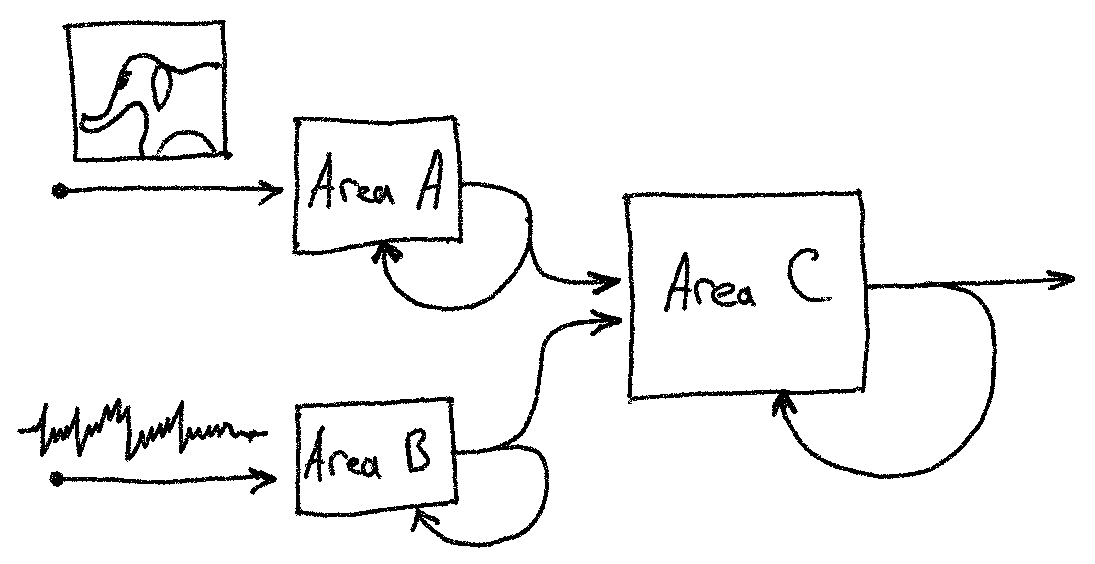
Area A encodes images, and area B - sound. The output of A and B is projected onto area C, which forms a combined image-sound representation. After several such projections (forward passes), the assemblies A-C and B-C become more and more overlapping - significantly more than by chance. This process is called association and described in 1. Following the example above, when areas A and B become associated, a sound an elephant makes will reconstruct a memory of elephant pictures (and vice versa), stored in B, assuming, of course, the presence of backward connections from area C to the incoming areas, which is not covered in this tutorial.
Input areas A and B can, of course, represent signals of the same modality that come from different cortical areas or layers.
Results¶
The example below shows area C activations, reshaped as matrices, when (from left to right):
only A is active;
only B is active;
both A and B are active.
before and after the association mechanism described above. Overlapping neurons are shown in green.
Before:

After:

More results¶
More results are here: http://85.217.171.57:8097. Pick “2020.11.26 AreaSequential assemblies” experiment from the drop-down list.
References¶
- 1(1,2)
Papadimitriou, C. H., Vempala, S. S., Mitropolsky, D., Collins, M., & Maass, W. (2020). Brain computation by assemblies of neurons. Proceedings of the National Academy of Sciences.
- 2
Willshaw, D. J., Buneman, O. P., & Longuet-Higgins, H. C. (1969). Non-holographic associative memory. Nature, 222(5197), 960-962.